






En aquest primer article dedicat a la millora genètica i la biotecnologia volem començar fent un repàs als diversos tipus de marcadors que s’utilitzen en els programes de millora genètica, des d’aquells visuals i/o morfològics usats en la millora clàssica fins els marcadors moleculars utilitzats per fer una selecció precoç. També es tracta el seu ús en la construcció de mapes genètics i les diferents aproximacions pel mapeig de caràcters d’interès.
La millora genètica va començar fa més de 8000 anys amb l’inici de l’agricultura sedentària i la domesticació dels primers cultius, però no va ser fins a principis del segle XX quan es va produir el seu enlairament amb el redescobriment dels treballs de Mendel de finals del segle XIX que determinen les lleis de l’herència. En l’actualitat és una disciplina molt amplia degut als progressos de la biologia i la genètica molecular, desenvolupant-se noves tècniques basades en la seqüència de l’ADN.
La millora genètica vegetal es defineix com l’elecció feta per l’home de les millors plantes escollides dins d’una població en la que existeix variabilitat. Per tant, les premisses més importants pel plantejament d’un programa de millora són: l’existència de variabilitat, la capacitat de detectar-la i de manipular-la per produir una varietat més estable.
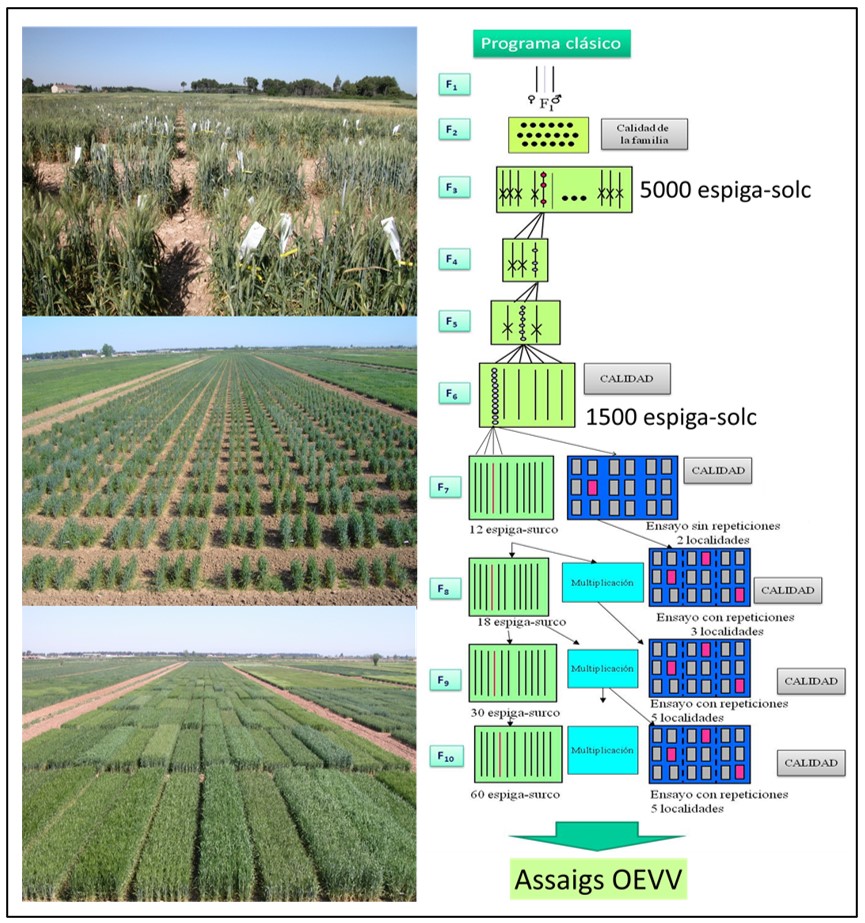
La millora genètica clàssica es basa en l’encreuament de les millors varietats i la selecció de la descendència mitjançant caràcters morfològics o visuals com la resistència a malalties i rendiment, els denominats marcadors morfològics. Aquest procés pot tardar fins a deu-onze anys en el cas de cereals o fins a mig segle en el cas de fruiters. La figura 1 mostra de manera esquemàtica un programa de millora de blat, on durant els primers anys la selecció es fa en espiga-solc. En aquests assajos es tenen en compte caràcters relacionats amb la qualitat, resistència a malalties i components del rendiment (nombre d’espigues, nombre de grans per espiga, pes del gra). A partir del sisè any es comença a fer una selecció en micro-parcel·les per seleccionar les línies mes productives.
El principal problema de l’ús de marcadors morfològics es la seva alta influència ambiental i baixa repetibilitat. Durant la dècada dels anys 50 van aparèixer els primers marcadors bioquímics, però no va ser fins la dècada dels anys 90 quan es va donar un gran impuls a la millora genètica vegetal amb l’aparició de la biologia molecular, en particular de la PCR (per les sigles en anglès de ‘reacció en cadena de la polimerasa) i els marcadors basats en ADN (Figura 2). L’avantatge d’aquest tipus de marcadors es que poden detectar-se en qualsevol estadi de desenvolupament de la planta, no estan influenciats per les condicions ambientals i es poden trobar en un nombre pràcticament il·limitat. A partir d’ells s’han pogut desenvolupar mapes genètics i localitzar els gens responsables d’alguns caràcters.
Els primer tipus de marcadors d’ADN que van sorgir van ser els RFLP, basats en hibridació amb sondes radioactives que feia molt laboriós el seu maneig. Per solucionar aquesta situació es van desenvolupar marcadors basats en la PCR com els RAPD, AFLP o SSR. Els dos primers tipus es van abandonar progressivament degut a la seva aleatorietat i baixa repetibilitat. Pel contrari, els SSR s’han utilitzat durant molts anys per la seva robustesa, distribució al llarg del genoma i la gran variabilitat que tenen. No ha sigut fins el darrers anys que han començat a substituir-se pels SNP. Aquests marcadors es caracteritzen per presentar polimorfisme en una sola base de l’ADN, el que fa que estiguin molt representats als genomes. El ràpid avenç en la tecnologia de genotipat d’alt rendiment on es poden obtenir milers de marcadors ha rebaixat molt el preu per genotipar una varietat, fent que avui els SNP siguin els marcadors elegits per la majoria de grups de recerca.

Figura 2. Amplificació mitjançant la PCR. A: aparell de PCR; B: cubeta d’electroforesi en gel d’agarosa; C: resultats de PCR en gel d’agarosa visualitzat sota raigs UV.
Aquesta nova tecnologia permet la construcció de mapes genètics altament saturats i, conjuntament amb la seqüenciació completa del genoma, una identificació de gens candidats molt ràpida i econòmica.
El marcadors moleculars s’han usat àmpliament per la construcció de mapes genètics. Aquests representen la posició dels marcadors moleculars al llarg dels cromosomes i es basen en els esdeveniments de recombinació entre els individus de la població de mapeig (Figura 3). Les diferents aplicacions dels mapes genètics en la millora vegetal son la identificació de les associacions entre els marcadors moleculars i els caràcters d’interès pel descobriment dels gens responsables dels caràcters. També s’utilitzen per la comparació de genomes entre diferents especies i el mapeig físic per facilitar l’acoblament dels genomes.
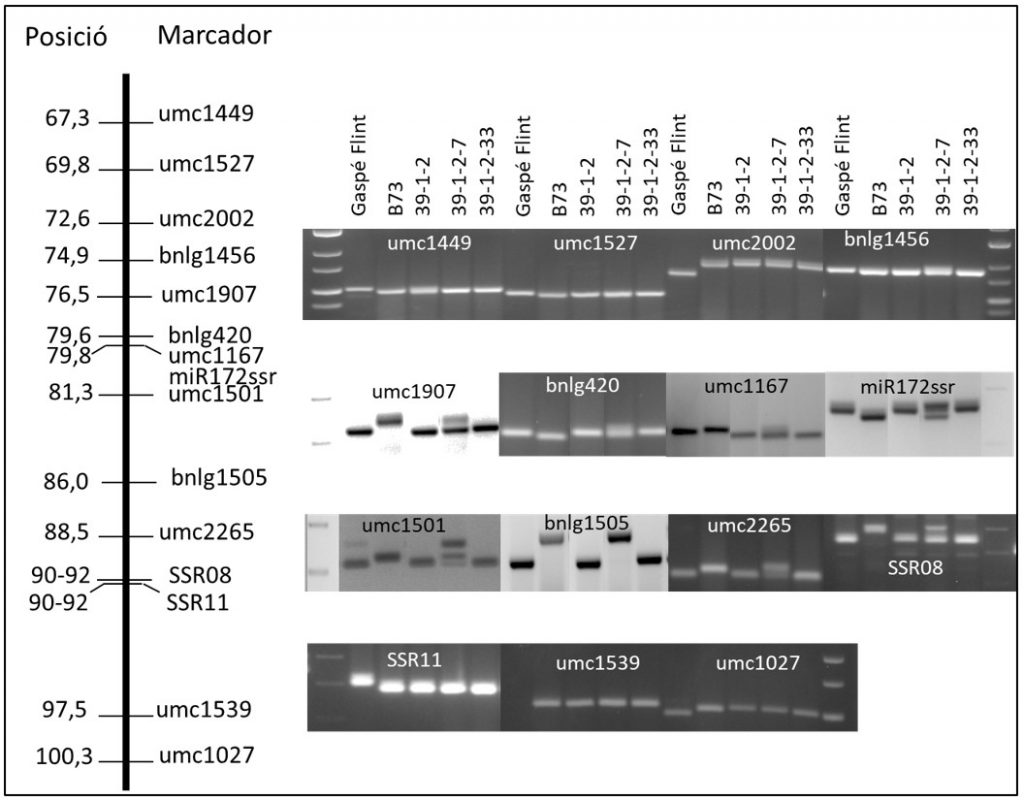
Figura 3. Representació de la part central del cromosoma 3 de panís i polimorfisme dels marcadors en la població resultant de l’encreuament B73 x Gaspé Flint.
Els caràcters a mapejar es poden dividir en simples i complexos. Un caràcter simple es aquell controlat per un gen, mentre que els caràcters complexos estan controlats per dos o més gens, els denominats QTL per les sigles en anglès ‘quantitative trait loci’. Els caràcters simples presenten una herència mendeliana i són mapejats com un marcador més, fent molt fàcil la seva localització cromosòmica. Pel contrari, els caràcters complexos presenten diferents tipus d’herència o efectes (dominant, additiva, epístasi, pliotropia) i pel seu mapeig és necessari l’ús de programes específics.
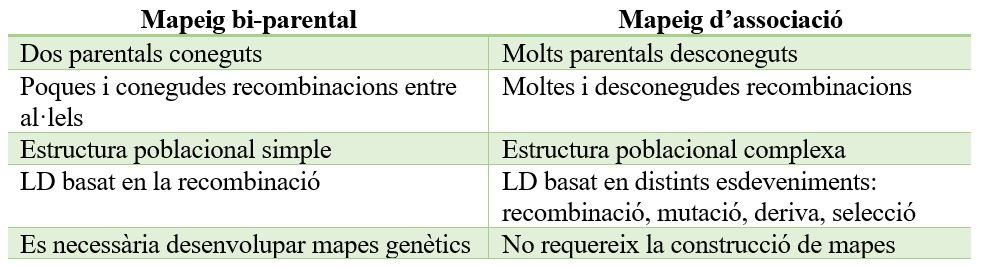
Clàssicament, el mapeig de QTL s’ha realitzat en poblacions derivades d’un encreuament entre dos parentals que presenten diverses característiques fenotípiques, el denominat mapeig bi-parental o mapeig de lligament. L’èxit del mapeig depèn de la densitat de marcadors que hi ha al mapa genètic, el nombre d’individus de la població i l’heretabilitat del caràcter. Per solucionar les limitacions d’aquest mapeig bi-parental va sorgir un altre tipus de mapeig, l’anomenat mapeig per associació o GWAS (per les sigles en anglès, genome wide association study). Aquest és una aproximació complementaria que proporciona una cobertura d’al·lels més amplia i una major resolució del mapeig. Les diferències entre ambdues metodologies es troben a la Taula 1. El GWAS es basa, al contrari que el mapeig bi-parental clàssic, en el desequilibri de lligament (LD), i es defineix com l’associació no feta a ‘atzar d’al·lels de diferents gens. Per l’anàlisi GWAS, enlloc de poblacions derivades de dues parentals, s’utilitzen col·leccions de varietats amb moltes i desconegudes relacions entre elles, pel que es fa necessari fer un anàlisi previ de la seva diversitat genètica.
Taula 1. Comparació del mapeig de QTL bi-parental i GWAS
Els mètodes de selecció dels programes de millora han evolucionat al llarg del temps. La millora clàssica es basava en la selecció fenotípica, que implica la selecció de les varietats amb els caràcters d’interès usant característiques visuals o morfològiques com el rendiment, data de floració, malalties. El principal inconvenient d’aquest tipus de selecció és el temps invertit i el nombre de persones necessàries per a fer-la.
Els marcadors moleculars són una alternativa per incrementar l’eficàcia dels programes de millora mitjançant la selecció assistida per marcadors o MAS (Figura 4). Aquesta permet fer una selecció primerenca dels individus i, per tant, reduir la mida de la població a avaluar simplificant el procés de selecció. La MAS depèn del lligament genètic entre el caràcter i el marcador molecular i, per tant, és necessari conèixer la seva posició al mapa genètic per escollir aquells marcadors més propers al caràcter. Aquesta selecció es pot fer únicament amb caràcters controlats per un o molt pocs gens. La selecció assistida s’ha emprat principalment en la millora de la resistència a malalties.
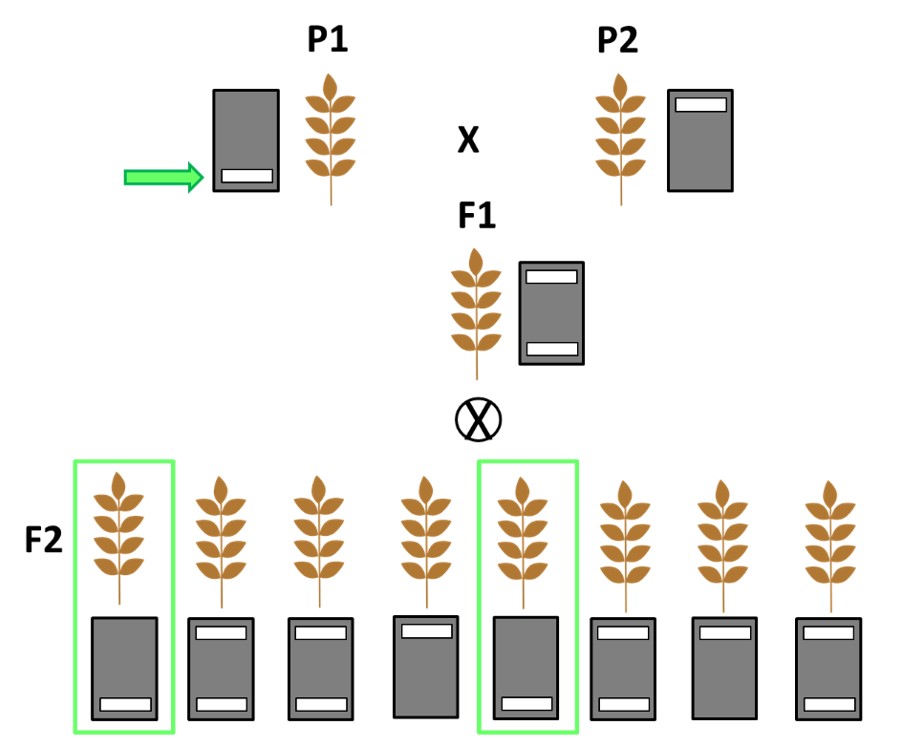
Figura 4. Esquema de selecció assistida en un programa de millora amb línies pures. L’encreuament entre dos línies parentals amb dotació al·lèlica diferent origina una F1 amb tos els individus heterozigots. L’autopol·linització d’aquesta F1 genera una F2 amb segregació. Solament els individus amb la dotació al·lèlica d’interès en homozigosis es seleccionen per als darrers encreuaments, eliminant el 75% de les plantes.
Jose Miguel Soriano
Rubén Rufo
Programa de Cultius Extensius Sostenibles, IRTA, Lleida











